


Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
Initialising ...
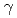 線照射によるイネ種子蛋白質組成突然変異体の効率的選抜
線照射によるイネ種子蛋白質組成突然変異体の効率的選抜西村 実*; 森田 竜平*; 草場 信*; 山口 博康*; 長谷 純宏
no journal, ,
イネの種子蛋白質突然変異体をSDS-PAGEによってM2種子でスクリーニングする場合、突然変異体の出現率は非常に低く、多大な労力が必要である。今回、玄米の外観に関して一次スクリーニングを行い、それらの中から種子蛋白質組成に関して2次スクリーニングを行うことによって種子蛋白質突然変異体を効率的に獲得できることを明らかにした。線又は炭素イオンビームを照射したイネのM2世代において粉質変異を示した287系統のうち68系統(線31系統,イオンビーム37系統)について蛋白質組成に変異が認められた。多くは26kDaのグロブリンと16kDaのプロラミンが同時に減少するタイプ、26kDaのグロブリンと16kDaのプロラミンが同時に減少し、さらにグルテリンの酸性サブユニットの減少を伴うタイプ、57kDaのグルテリンの前駆体が増加するタイプの3種類であった。今後これらの変異形質が安定的に後代に伝わるか否か、致死性等の劣悪変異を伴っていないか等について確認を行う必要があるが、種子蛋白質組成の変異体のスクリーニングに非常に有効な選抜方法になり得ると考えられる。
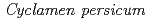
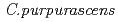 )の突然変異体作出
)の突然変異体作出近藤 恵美子*; 長谷 純宏; 田中 淳; 石坂 宏*
no journal, ,
シクラメンの栽培種(, 2n=2x=48)と芳香性野生(, 2n=2x=34)の種間交雑から雑種(2n=41)を育成し、その染色体倍加によって複二倍体(2n=4x=82)の芳香性品種を育成した。しかし、育成品種は花色と花形の変化が乏しく、形質の多様化が必要である。そこで培養細胞へのイオンビーム照射により、多様な変異体の作出を試みた。 と
と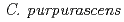 の複二倍体(2n=82)数系統の花弁,培養中の黄化葉柄,花糸由来の不定胚を供試し、花弁には220MeV炭素イオンを、黄化葉柄及び不定胚には320MeV炭素イオンをそれぞれ0-50Gyで照射した。生存率,塊茎重並びに不定芽形成率から判断して、花弁,黄化葉柄,不定胚すべての組織で1-2Gyの線量が適当と推察された。春までに開花した個体について形態観察を行った。花弁では、花色の濃淡,花色変異,斑入り,花弁の切れ込み(フリンジ),花の大小,奇形等の変異が見られ、葉では斑入りやアントシアニンの沈着が、花梗は太さや長さに変異が認められた。変異体獲得頻度と変異形質の発現には系統間,供試部位により差が認められた。
の複二倍体(2n=82)数系統の花弁,培養中の黄化葉柄,花糸由来の不定胚を供試し、花弁には220MeV炭素イオンを、黄化葉柄及び不定胚には320MeV炭素イオンをそれぞれ0-50Gyで照射した。生存率,塊茎重並びに不定芽形成率から判断して、花弁,黄化葉柄,不定胚すべての組織で1-2Gyの線量が適当と推察された。春までに開花した個体について形態観察を行った。花弁では、花色の濃淡,花色変異,斑入り,花弁の切れ込み(フリンジ),花の大小,奇形等の変異が見られ、葉では斑入りやアントシアニンの沈着が、花梗は太さや長さに変異が認められた。変異体獲得頻度と変異形質の発現には系統間,供試部位により差が認められた。
長谷 純宏
no journal, ,
本講演では、花弁形態に関する変異体(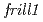 )及び紫外線耐性変異体(
)及び紫外線耐性変異体(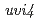 )の解析から明らかになった新たな植物機能について紹介する。は、花弁とガクの周縁がフリル状に変化する突然変異体である。この変異体では、花弁発達の後半において花弁先端部の細胞の配列が乱れ、異所的な核内倍加が起きていることがわかった。変異体では、
)の解析から明らかになった新たな植物機能について紹介する。は、花弁とガクの周縁がフリル状に変化する突然変異体である。この変異体では、花弁発達の後半において花弁先端部の細胞の配列が乱れ、異所的な核内倍加が起きていることがわかった。変異体では、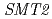 遺伝子に1塩基の欠失が生じており、異常なステロールの成分比により通常では核内倍加が起こらない組織で核内倍加が起きたものと考えられた。一方、は、UV-B環境下で野生型に比べて生育が抑制されにくい変異体である。では、機能未知の塩基性タンパクをコードするAt2g42260遺伝子に123 bpの欠失があることがわかった。各組織の倍数性を調査したところ、野生型よりも核内倍加が促進されており、細胞の倍数性が上昇することで紫外線耐性になっていることが示唆された。以上の結果から、核内倍加を抑制することがArabidopsisの花弁の形態形成に必要であり、一方、核内倍加を促進することは劣悪環境に耐えて効率よく組織を肥大化させる戦略であると考えられた。
遺伝子に1塩基の欠失が生じており、異常なステロールの成分比により通常では核内倍加が起こらない組織で核内倍加が起きたものと考えられた。一方、は、UV-B環境下で野生型に比べて生育が抑制されにくい変異体である。では、機能未知の塩基性タンパクをコードするAt2g42260遺伝子に123 bpの欠失があることがわかった。各組織の倍数性を調査したところ、野生型よりも核内倍加が促進されており、細胞の倍数性が上昇することで紫外線耐性になっていることが示唆された。以上の結果から、核内倍加を抑制することがArabidopsisの花弁の形態形成に必要であり、一方、核内倍加を促進することは劣悪環境に耐えて効率よく組織を肥大化させる戦略であると考えられた。